Какой расход бензина у Лада Гранта на 100 км, 8 и 16 клапанов
Содержание
- 1 Заводские параметры
- 1.1 Расход топлива 8-ми клапанных двигателей
- 1.2 Расход топлива 16-ти клапанных двигателей
- 2 Как измерить расход топлива самому?
- 2.1 Методика по контрольной лампе
- 2.2 От бака до бака
- 3 Основные мировые методики измерения расхода новых автомобилей
- 3.1 NEDC
- 3.2 FTP-75
- 3.3 Японский цикл JC08
- 4 Основные технические факторы, которые влияют на расход топлива
Заводские параметры
Существуют определённые методики для определения среднего расхода топлива двигателем автомобиля. Они кстати утверждены законодательно. Автомобиль не тестируют на полигонах, а пригоняют в специальные лаборатории с «идеальными» условиями. Далее устанавливают колёса машины на барабан, которые имитируют городской и загородный условия эксплуатации. Именно поэтому в большинстве случаев реальный расход не совпадает с заводским.
Добавим к этому ещё и тот фактор, что манера езды отличается у большинства водителей.
Факторы, влияющие на расход топлива
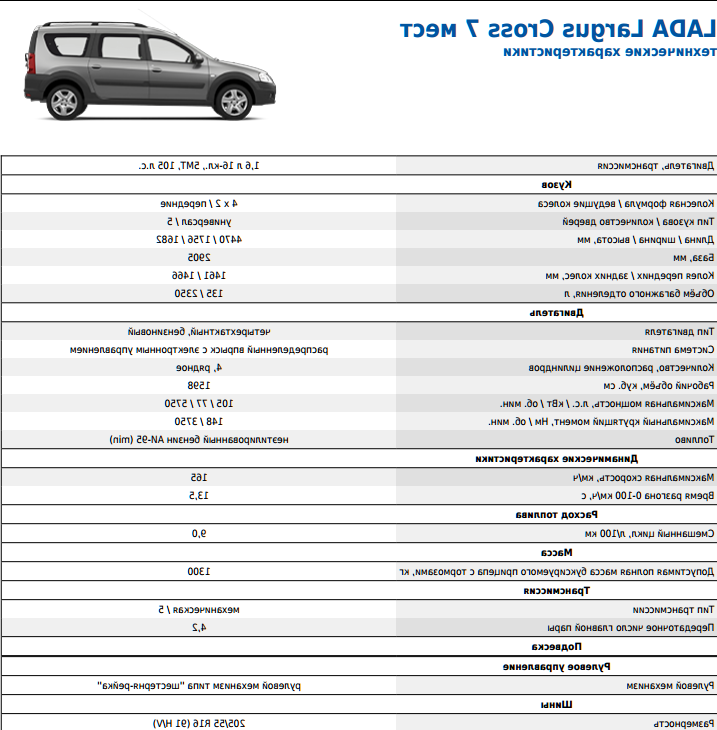
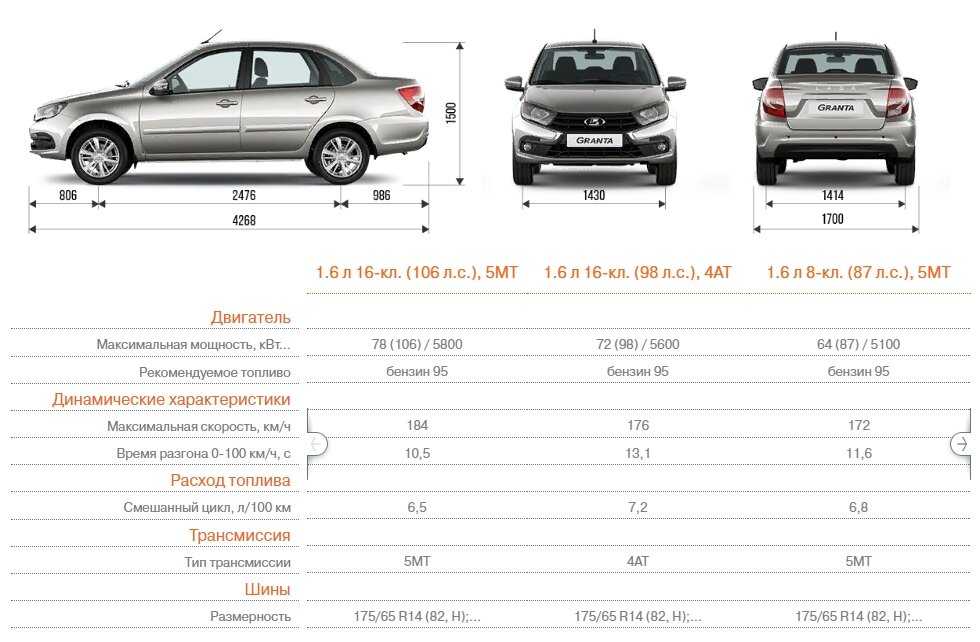
Расход топлива 8-ми клапанных двигателей
Данные представлены для автомобилей Лада Гранта оснащенных МКПП.
| Двигатель (модель) | Мощность двигателя (л.с.) | Объём двигателя (куб.см) | Расход топлива в городском цикле (л/100 км) | Расход топлива за городом (л/100 км) | Расход топлива в смешанном цикле (л/100 км) |
|---|---|---|---|---|---|
| 11183 | 82 | 1596 | 9.7 | 6.1 | 7,4 |
| 11186 | 87 | 1596 | 9 | 5.8 | 7,0 |
| 21116 | 87 | 1596 | 9 | 5.8 | 7,0 |
Подробно о самом распространённом двигателе линейки Лада Гранта 21116 читайте здесь.
Расход топлива 16-ти клапанных двигателей
Данные представлены для автомобилей Лада Гранта оснащенных МКПП.
| Двигатель (модель) | Мощность двигателя (л.с.) | Объём двигателя (куб.см) | Расход топлива в городском цикле (л/100 км) | Расход топлива за городом (л/100 км) | Расход топлива в смешанном цикле (л/100 км) |
|---|---|---|---|---|---|
| 21126 | 98 | 1596 | 9.9 | 6.1 | 7,6 |
| 21127 | 106 | 1596 | 8.6 | 5.6 | 6,7 |
Как видно из таблиц расход 16-ти клапанных двигателей чуть ниже 8-ми клапанных. А стоит версия с мощным двигателем дороже. Поэтому мы хотим спросить Ваше мнение, какой двигатель лучше.
Все испытания проводятся на 95-ом бензине, именно под это октановое число рассчитаны все двигатели Лада Гранта.
Как измерить расход топлива самому?
Существуют два основных способа измерения среднего расхода топлива. В случае, если расход топлива Вашим автомобилем будет существенно отличаться от заводских параметров, то необходимо диагностировать причину.
В случае, если расход топлива Вашим автомобилем будет существенно отличаться от заводских параметров, то необходимо диагностировать причину.
Методика по контрольной лампе
Самый простой из них, и как мне кажется самый необъективный, осуществляется по контрольной лампе топлива. Суть его состоит в следующем:
- как только у Вас загорелась контрольная лампа «бензоколонки», Вам необходимо заправить 20 литров топлива.
- Обнулить суточный пробег.
- Дождаться пока загорится снова контрольная лампа «бензоколонки» и поделить пройденный путь на 2. К примеру, если Вы проехали 300 км, то необходимо поделить 300 на 2 (300/2=150 км). Значит на 10 литрах Вы проезжаете 150 км. Или поделить 20 на 300 и умножить на 100, получится, что в данном случае Ваш расход топлива составил 6,6 литра на сто км пути.
На 20 литрах автомобиль проехал 412 км
Этот способ нельзя назвать точным, особенно учитывая то, что контрольная лампа на Лада Гранта загорается когда ей захочется, от любой горки, может загореться даже от резкого нажатия на педаль тормоза (экстренное торможение).
От бака до бака
Этот способ самый точный, если соблюдать все правила. Суть его заключается в следующем:
- при минимальном остатке топлива в баке, необходимо залить полный бак, до первого «отстрела» пистолета бензоколонки.
- Обнулить суточный пробег.
- Дождаться пока уровень топлива упадёт до минимальной отметки (чем меньше уровень, тем точнее будет измерение) и заправиться до полного бака снова, до первого «отстрела» пистолета бензоколонки.
- Далее сопоставить показания суточного пробега и количества топлива, которое поместилось в бензобак. Например, если Вы проехали 500 км, а потратили 41 литр топлива, то расход топлива автомобилем составит (41/500)*100= 8,2 (8,2 литра на 100 км.)
Именно по этому способу я рекомендую сверять показания бортового компьютера и реального расхода топлива Вашим автомобилем!
Средний расход топлива по бортовому компьютеру
Основные мировые методики измерения расхода новых автомобилей
Как мы писали выше, существует несколько методик по измерению среднего расхода топлива у автомобилей. Расскажем о них подробнее.
Расскажем о них подробнее.
NEDC
На территории Европы чаще всего применяется цикл типа NEDC, который был разработан ещё около сорока лет тому назад. Это и вызывает упрёки со стороны экспертов, которые говорят о несовременности метода.
Структура алгоритма NEDC
Главными особенностями можно считать то, что производителям позволено использовать селективный метод выбора машины. Мы говорим о том, что для тестирования предоставляются авто с уже обкатанными узлами, специальными смазочными материалами пониженной вязкости, а также с наиболее компактными шинами, которые могут устанавливаться на автомобиль.
Помимо этого, при желании, тестирование может проходить с выключенным радио, обогревом и так далее. Это позволяет «подобрать» те результаты, которые бы наилучшим образом подошли производителю.
Измерения проводятся в городском (NEDC UDC) и загородном (NEDC EUDC) цикле. Длина «городского теста» составляет 780 секунд, тогда как загородное движение предполагает период времени, равный 400 секундам. В первом случае средняя скорость будет немногим более 18 километров, а во втором — около 62,6 километров в час.
В первом случае средняя скорость будет немногим более 18 километров, а во втором — около 62,6 километров в час.
Критикуется данная методика по той причине, что многие модели с дополнительным электрическим мотором могут проехать большую часть этой короткой дистанции, практически не используя двигатель внутреннего сгорания. Это и становится источником удивительных результатов с крайне малым расходом топлива, даже спортивными авто.
FTP-75
Но эти стандарты уже не удовлетворяют требованиям рынка, так что производители чаще обращаются к американскому стандарту FTP-75. Он считается наиболее приближенным к реальности, хотя эксперты говорят о проблемах с отсутствием участков, на которых движение происходит равномерно.
Зависимость скорости алгоритма FTP-75
Он похож на европейский NEDC, включая и среднюю скорость, равную 34,1 километрам в час. Преимуществом можно считать и то, что длительность испытаний составляет 1874 секунды, так что гибридным моделям автомобилей сложнее показывать «нереально низкие» показатели расхода топлива. Но, что самое главное, так это то, что американская версия теста обязует производителей включать кондиционер, если он предусмотрен в комплектации.
Но, что самое главное, так это то, что американская версия теста обязует производителей включать кондиционер, если он предусмотрен в комплектации.
Как результат, расход топлива по этому тесту оказывается выше, чем в NEDC на 10 или даже 20 процентов.
Японский цикл JC08
В случае с JC08 разница будет ещё большей. Дело в том, что в нём количество разгонов и торможений является максимальным, а длительных участков с ровными скоростными параметрами нет вовсе.
Кроме того, максимальная скорость не превышает отметки в 80 километров в час. На одном из участков, длина которого равна 8,2 километрам, средняя скорость не достигает даже 25 километров в час.
Cамая «медленная» из известных методик расчёта среднего расхода
Конечно же, можно говорить о жёсткости теста, но он учитывает повышенную загрузку улиц, которая характерна для островной страны Японии. К тому же, в таких условиях пребывают водители большинства больших городов по всему миру. Так что, стоит рассматривать данную методику, как наиболее приближенную к реальности.
Так что, стоит рассматривать данную методику, как наиболее приближенную к реальности.
В общем же, оптимального метода тестирования пока не существует. Существующие варианты представляют собой компромисс, который автомобильные производители стараются использовать себе на пользу. Поэтому нужно всегда стараться учитывать конкретную методику, которая дала результаты, указанные в спецификациях автомобиля.
Основные технические факторы, которые влияют на расход топлива
В основном на расход топлива автомобилем влияет:
- Качество самого топлива. Если Вам не повезло и вместо хорошего топлива автомобиль был заправлен «бадягой», то придётся сливать весь бензин из бака.
- чистота фильтров (воздушный и топливный фильтр). Данные поломки можно диагностировать самому. Наиболее распространённые причины — не соблюдение сроков замены фильтров.
- Правильная работа датчиков и системы управления двигателем.
- Не верная работа самого двигателя в результате износа или не правильной обкатки.

- Сдутые шины. Не верно подобранное давление в автомобильной шине сильно влияет на расход топлива.
- Не верно подобранный тип моторного масла. Чтобы этого не произошло необходимо обращать тщательное внимание на выбор масла. Подробнее в материалах: о выборе масла в 8-ми клапанный двигатель, о выборе масла в 16-ти клапанный двигатель.
- Износ тормозных механизмов. В данном случае возможнен контакт колодки с тормозным диском в результате поломки тормозного механизма. Данная ситуация очень распространена. Виной этому плохие водосливы, вода попадает в тормозные механизмы и нарушает их работу, выбивает оттуда смазку, в результате чего происходит «закисание» механизмов.
- Движение на непрогетом двигателе. Это конечно спорный вопрос. Из-за особенности конструкции охлаждающей системы температура двигателя редко в холодную погоду поднимается выше 80 градусов. Раньше данное явление считалось поломкой, теперь, наоборот, АвтоВаз выпустил обратное предписание и убрал на первых моделях индикатор температуры двигателя вовсе.

Расход топлива Лада Гранта на 100 км: отзывы владельцев
Содержание статьи
- Lada Granta с мотором 1.6 82 л.с.
- Отзывы о расходе топлива
- Еще отзывы
- Lada Granta с двигателем на 87 л.с.
- Отзывы владельцев о расходе топлива
- Еще отзывы
- Lada Granta с мотором 1.6 на 98 сил
- Показатели реального расхода по отзывам
- Еще отзывы
- Характеристики Lada Granta с мотором 1.6 на 106 сил
- Отзывы о расходе от реальных владельцев
- Еще отзывы
Модель Лада Гранта появилась на рынке в 2011, где была призвана заменить Лада Калина. Машина сразу привлекла внимание новшествами и широким выбором комплектаций – базовой версией Стандарт, а также вариантами Норма и Люкс. Посмотрим, какой расход топлива Лада Гранта на 100 км пути.
Автомобиль имеет целый набор преимуществ:
- Несколько модификаций 8-ми и 16-ти клапанных бензиновых моторов 1. 6 – надежных и мощных агрегатов.
- Надежная трансмиссия, появление первого ВАЗовского автомата.
- Адаптированная к отечественным дорогам и климаты подвеска.
- Экономичный расход топлива всех версий Лада Гранта.
- Приемлемая цена в любой из комплектаций.
 6 – надежных и мощных агрегатов.
6 – надежных и мощных агрегатов.Lada Granta с мотором 1.6 82 л.с.
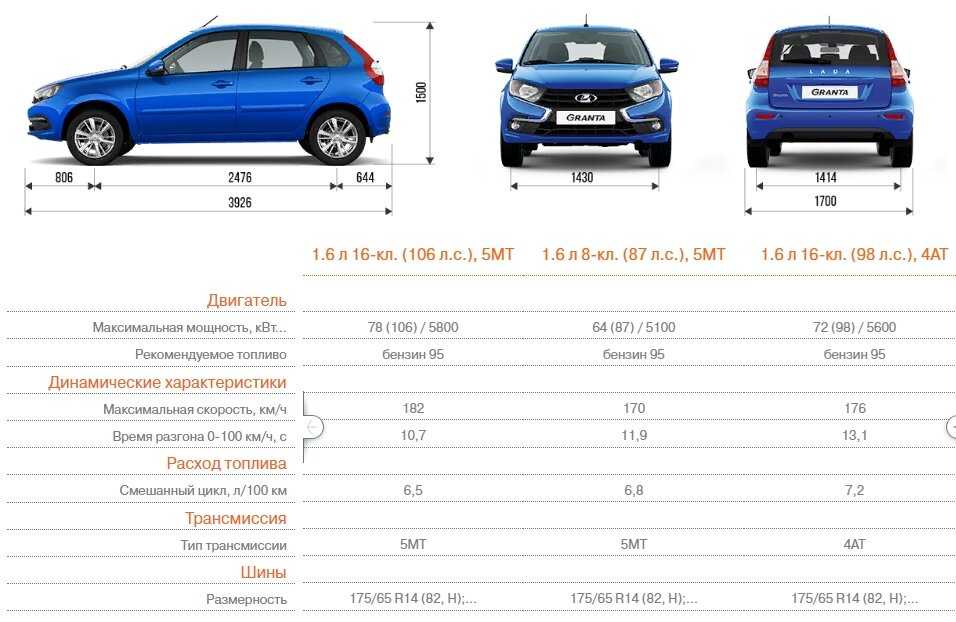
В самой доступной версии Lada Granta оснащается бензиновым восьмиклапанным силовым агрегатом 1.6 с инжектором. Он развивает 82 лошадиных силы при моменте сил 132 Н·м и разгоняет машину до сотни за 12,6 с при максималке 165 км/ч. Расход топлива Лада Гранта на 100 км на этой комплектации:
- город 9,7 л;
- смешанный 7,4 л;
- трасса 6,1 л.
Отзывы о расходе топлива
- Антон, Севастополь. Владел подержанной иномаркой, а в 2013 взял Лада Гранта. Новый автомобиль оправдал ожидания – симпатичный седан, отлично чувствующий себя в городе, с неплохими возможностями на грунтовой дороге благодаря адаптированной подвеске. Моторчик на 82 силы неплохо тянет, уже прошел 100 тыс. км – никаких проблем, только масло и маслофильтр менял. По городу при включенном кондиционере расход от 10 л в загородном цикле можно уложиться в расход топлива Лада Гранта на 100 км — 6 литров.
- Валерий, Рязань. У меня Лада Гранта с объемом двигателя 1.6 и мощностью 82 лошади, купил в 2014. До этого у меня было много ВАЗовских моделей, эту считаю наиболее сбалансированной, надежной и комфортной. При желании недостатки найти можно, но цена полностью соответствует предлагаемому качеству. Очень надежная ходовка, пока менял только тормозные колодки. В смешанном цикле реальны расход топлива около 7,5 л на сотню, заправляюсь АИ 95.
- Богдан, Ставропольский край. Свою Лада Гранта купил в 2015 году, пока не жалуюсь. Жесткая подвеска очень вынослива и это меня устраивает, мотор на 82 силы с системой распределенного впрыска топлива отлично справляется со своими обязанностями. Единственное, что настораживает, движок берет масло, расход бензина у него при этом не более 10 литров даже зимой.
 Моторчик на 82 силы неплохо тянет, уже прошел 100 тыс. км – никаких проблем, только масло и маслофильтр менял. По городу при включенном кондиционере расход от 10 л в загородном цикле можно уложиться в расход топлива Лада Гранта на 100 км — 6 литров.
Моторчик на 82 силы неплохо тянет, уже прошел 100 тыс. км – никаких проблем, только масло и маслофильтр менял. По городу при включенном кондиционере расход от 10 л в загородном цикле можно уложиться в расход топлива Лада Гранта на 100 км — 6 литров.
Еще отзывы
- Денис, Брянск. У меня Лада Гранта, модель с мощностью двигателя 82 силы с МКПП. Отличный вариант для города, один из самых надежных автомобилей ВАЗ. Отмечу надежность и приемлемый расход топлива Лада Гранта на 100 км в районе 7 л, зимой немного больше, чем летом. К недостаткам отнесу никакую шумоизоляцию, что утомляет при длительных поездках.
- Анатолий, Пермь. Мне требовался экономный недорогой автомобиль для города, поэтому остановил свой выбор на Лада Гранта с движком на 82 силы. Салон комфортный, но жесткий пластик создает акустический дискомфорт. К подвеске, движку и трансмиссии претензий нет. Машина потребляет 6 л на трассе и до 9 по городу.
- Сергей, Харьков. У меня Lada Granta 2012 года с мотором 1.6 на 82 лошадиных сил и механической коробкой. За все время никаких проблем не возникло, салон как для Лады очень комфортен, багажник вместителен. Ходовая жестковата, но очень надежна, по двигателю менял только масло и фильтра. Средний расход составляет около 7 л АИ 95.
 Средний расход составляет около 7 л АИ 95.
Средний расход составляет около 7 л АИ 95.Lada Granta с двигателем на 87 л.с.
Более мощная модификация Лада Гранта оснащена мотором с тем же объемом 1.6 на 87 лошадиных сил с моментом 140 Н·м и улучшенной динамикой в 12,2 с до первой сотни. Максимальная скорость в этой комплектации составляет 167 км/ч, а расход топлива Лада Гранта на 100 км:
- в трафике 9 л;
- в среднем 7 л;
- на шоссе 5,7 л.
Отзывы владельцев о расходе топлива
- Александр, Керчь. Брал Лада Гранта в 2016 с салона, поскольку мне всегда нравилась внешность автомобиля. Выбрал вариант с восьмиклапанным двигателем 1.6 на 87 лошадей, поскольку считаю его самым надежным. Поставил машину на обслуживание, пока менял только масло и фильтры. Что касается расхода топлива – на шоссе получается 6 л, по городу получается не более 10 даже зимой.
- Степан, Самара. Для моей семьи из четырех человек Lada Granta – лучший вариант. Она хорошо чувствует себя в городе, на ней можно вывести семью на дачу или пикник. В багажнике можно разместить все, что нужно. Подвеска и трансмиссия надежны, мотор на 87 сил отлично тянет, никаких проблем с ним не было. Средний расход топлива Лада Гранта на 100 км 7,5 л летом и около 8 зимой.
- Ярослав, Петербург. Покупал Лада Гранта в салоне, надеясь на высокое качество сборки, которое декларировал производитель. Может, мне досталась такая машина, но я ней недоволен, особенно в плане надежности и комфорта. Салон шумный, постоянные проблемы с подвеской и системой подачи топлива. Расход, правда небольшой, 6-9 л на сотню, но все равно думаю продавать ее.
 Что касается расхода топлива – на шоссе получается 6 л, по городу получается не более 10 даже зимой.
Что касается расхода топлива – на шоссе получается 6 л, по городу получается не более 10 даже зимой.Еще отзывы
Сколько ест Лада Гранта?
- Иван, Оренбург. У меня Лада Гранта 1.6 2104 года с механической коробкой. Пока все нравится, машина надежная, неприхотливая, запчасти недорогие. Подвеска адаптирована к нашим дорогам, двигатель Лады Гранты на 87 л.с. легко заводится в самые лютые морозы. Расход топлива по трассе 6 л, в пробках повышается до 9.
- Вячеслав, Ханты-Мансийск. Пробрел Лада Гранта в 2015, брал в салоне, с мотором на 87 лошадей. Был приятно удивлен качеством сборки, комфортом в салоне, совершенно не характерным для отечественного автопрома. Подвеска несколько жестковата, но отлично разбирается с дорожными неровностями. Расход топлива не превышает 9 л даже зимой, на трассе можно уложиться в 6 л.
- Николай, Ростов. Брал свою Лада Гранта в 2016 с рук, мотор 1.6 на 87 сил с пятиступенчатой механикой. Пока проехал менее 20 тыс. км, никаких претензий к машине не имею. Она легко управляется, динамична, дешева в ремонте. Просторный и удобный салон, багажник – все для людей. Расход тоже радует – по кругу 7 л, лью только АИ 95 или АИ 98.
 Пока все нравится, машина надежная, неприхотливая, запчасти недорогие. Подвеска адаптирована к нашим дорогам, двигатель Лады Гранты на 87 л.с. легко заводится в самые лютые морозы. Расход топлива по трассе 6 л, в пробках повышается до 9.
Пока все нравится, машина надежная, неприхотливая, запчасти недорогие. Подвеска адаптирована к нашим дорогам, двигатель Лады Гранты на 87 л.с. легко заводится в самые лютые морозы. Расход топлива по трассе 6 л, в пробках повышается до 9.Lada Granta с мотором 1.6 на 98 сил
Модификация Лада Гранта с шестнадцатиклапанным мотором 1. 6, на обладает повышенной мощностью в 98 сил и крутящим моментом 145 Н·м. На нее устанавливается четырехступенчатый автомат, что вывело этот автомобиль на новый уровень комфорта. При этом динамика авто немного снизилась до 13,3 с до сотни, а показатель максимальной скорости вырос до 173 км/ч. Расход топлива Лада Гранта на 100 км у этой версии составляет:
6, на обладает повышенной мощностью в 98 сил и крутящим моментом 145 Н·м. На нее устанавливается четырехступенчатый автомат, что вывело этот автомобиль на новый уровень комфорта. При этом динамика авто немного снизилась до 13,3 с до сотни, а показатель максимальной скорости вырос до 173 км/ч. Расход топлива Лада Гранта на 100 км у этой версии составляет:
- на загруженных улицах 9,9 л;
- средний показатель 7,6 л;
- за городом 6,1 л.
Показатели реального расхода по отзывам
- Василий, Екатеринбург. Лада Гранта выпуска 2015 года с двигателем 1.6 на 98 лошадей с АКПП. Пока машина заслуживает только хороших слов, радует работа автомата, вообще сборка просто удивительная, как для наших машин. Ходовая часть очень надежная, двигатель хорошо тянет, расход в от 7 до 10 литров, что меня вполне устраивает.
- Анатолий, Киев. Брал свою Ладу Гранту в салоне, для езды по городу выбрал версию с двигателем 1.6 и автоматической трансмиссией. Считаю это лучшим вариантом по соотношению цены и качества, при этом надежным и неприхотливым. Подвеска идеально адаптирована под наши дороги, при этом отлично держит поверхность, машина легкой управляется и уверенно тормозит двигателем. Наибольший расход бензина зимой в пробках не более 10,5 л, на трассе можно легко уложиться в расход топлива Лада Гранта на 100 км — 6 л АИ 95 или 98.
- Ильхан, Пятигорск. Лада Гранта – мой первый автомобиль, подарили родители. Версия 1.6 с АКПП и мощностью в 98 лошадей – идеальный вариант для начинающего автолюбителя. Машинка идеальна в городе, для поездки на дачу или на природу. Универсальная подвеска хорошо держит машину на трассе, не пробивается на ухабах, обеспечивает адекватную управляемость. Расход бензина не превышает 10 л, на чистой дороге получается около 7 л даже на скоростях свыше 120 км/ч.
 Подвеска идеально адаптирована под наши дороги, при этом отлично держит поверхность, машина легкой управляется и уверенно тормозит двигателем. Наибольший расход бензина зимой в пробках не более 10,5 л, на трассе можно легко уложиться в расход топлива Лада Гранта на 100 км — 6 л АИ 95 или 98.
Подвеска идеально адаптирована под наши дороги, при этом отлично держит поверхность, машина легкой управляется и уверенно тормозит двигателем. Наибольший расход бензина зимой в пробках не более 10,5 л, на трассе можно легко уложиться в расход топлива Лада Гранта на 100 км — 6 л АИ 95 или 98.Еще отзывы
- Дмитрий, Липецк. Как только Лада Гранта появилась в салонах, я сразу захотел получить это автомобиль. Выбрал версию с движком 1.6 и четырехступенчатым автоматом, хотя сомневался в его надежности. Но мои опасения оказались напрасными, машина идет отлично, без рывков, по надежности нет никаких нареканий. В остальном добротный автомобиль, отвечающий моим запросам по бензину 9,5 в городе на трассе расход топлива Лада Гранта на 100 км — 6 л, думаю поставить газовую установку.
- Григорий, Феодосия. Уже несколько лет владею Лада Гранта с 98-ильным движком и автоматом. В основном машиной доволен, единственная серьезная претензия – никакая шумоизоляция. Зато технические характеристики на уровне, приятно удивляет качество автоматической трансмиссии – идет плавно, передачи переключаются быстро и вовремя. Расход топлива на трассе около 6 л, в на городских улицах около 9.
- Максим, Серпухов. Все время ездил на б/у иномарках, а в 2015 решил взять машину с нуля – Лада Гранта. Должен сказать, что не пожалел о своем выборе. Динамичная, идеальная для городских условий машинка. Благодаря упругой и надежной подвеске она подходит и для поездок на дачу. Благо, багажник позволяет загрузить что угодно, в разумных пределах. Расход бензина Лады Гранты АИ 95 около 5,5 на трассе, в городе получается до 10 л зимой.
 В остальном добротный автомобиль, отвечающий моим запросам по бензину 9,5 в городе на трассе расход топлива Лада Гранта на 100 км — 6 л, думаю поставить газовую установку.
В остальном добротный автомобиль, отвечающий моим запросам по бензину 9,5 в городе на трассе расход топлива Лада Гранта на 100 км — 6 л, думаю поставить газовую установку.
Характеристики Lada Granta с мотором 1.6 на 106 сил
Самая мощная Лада Гранта комплектуется 16-ти клапанным бензиновым мотором на 106 сил с крутящим моментом 148 Н·м и динамикой до сотни за 10,9 секунды. Машина развивает максимальную скорость 183 км/ч. Расход топлива Лада Гранта на 100 км в этом варианте комплектации:
- в городском режиме 8,6 л;
- смешанный цикл 6,7 л;
- на шоссе 5,6 л.
Варианты с роботизированной коробкой практически не отличаются потреблением, но стоят дороже, поэтому не слишком популярны у отечественных автолюбителей.
Предлагается и спортивная версия с двигателем на 118 сил и крутящим моментом 154 Н·м за это приходится платить увеличением расхода топлива на Лада Гранта Спорт на 1 л в каждом из режимов движения.
Отзывы о расходе от реальных владельцев
- Игнат, Москва. Брал Ладу Гранту специально для поездок по городу и выбрал самый приемистый мотор на 106 сил с пятиступенчатой механикой. Очень удобный вариант – надежный двигатель и всеядная подвеска, плюс вместительный салон и багажник. Отлично чувствует себя в плотном потоке, ремонт пока не делал, только масло менял. Расход топлива Лада Гранта на 100 км по трассе 5,5 л, в городском потоке до 9 л.
- Петр, Магнитогорск. Брал Лада Гранта Люкс в 2015 с салона, модификацию, оснащенную роботом. Покупкой вполне доволен, боялся, что трансмиссия будет глючить. Но все нормально, не дергает, не перегревается, не ломается. Мотор на 106 сил отлично тянет как на перекрестках в городе, так и при обгонах на трассе. Салон достаточно комфортен, расход тоже радует и не превышает по кругу 7,5 л при такой динамике вполне пристойный показатель.
- Марат, Казань. Если брать машину с нуля, как это сделал я, Лада Гранта – лучший вариант по деньгам. За это я получил модель с движком 1.6 на 106 сил, выбрав версию с механикой, автомат побоялся брать. Сверчки в салоне убрал, сделал шумку – больше ничего не нужно. Ходовые качества вполне пристойные, комфорт стал на уровне, а расход от 5 до 9 л в разных режимах.
 Очень удобный вариант – надежный двигатель и всеядная подвеска, плюс вместительный салон и багажник. Отлично чувствует себя в плотном потоке, ремонт пока не делал, только масло менял. Расход топлива Лада Гранта на 100 км по трассе 5,5 л, в городском потоке до 9 л.
Очень удобный вариант – надежный двигатель и всеядная подвеска, плюс вместительный салон и багажник. Отлично чувствует себя в плотном потоке, ремонт пока не делал, только масло менял. Расход топлива Лада Гранта на 100 км по трассе 5,5 л, в городском потоке до 9 л.
Еще отзывы
Сколько ест Лада Гранта?
- Денис, Минск. Купил Лада Гранта в 2014, с тех пор почти не вылезаю из-за руля, проехал почти 200 тыс. км, просто машина нужна по работе. Должен заметить, что негативные отзывы по шумоизоляции не могу поддержать, у меня все в порядке, скрипов в салоне просто нету, может повезло со сборкой. Машина легко управляется, хорошо держит дорогу, 106 лошадиных сил хватает для уверенной езды по городу или трассе расход практически полностью соответствует прописанному в паспорте расход топлива Лада Гранта на 100 км от 6 до 9 л АИ 95.
- Алексей, Петрозаводск. Машину брал с салона, выбрал версию Лада Гранта на 106 лошадей с механикой. Пока машиной доволен, никаких проблем с поломками нету, моторчик тянет исправно, масло не берет пока новый. Езжу много как в городе, так и по трассе. Расход вполне адекватный от 5,5 до 10 л.
- Олег, Нижний Новгород. Машину взял в 2012 году выбрал версию на 106 сил на ручке. Считаю эту машину один из лучших творений отечественного автопрома. Подвеска жестковата, но отличается потрясающей надежностью. Мотор хорошо тянет в любой ситуации – на светофоре или обгоне фуры по шоссе. Клиренс позволяет без опаски съезжать с асфальта, поэтому часто езжу на дачу, вывожу семью на природу. Что касается потребления, то расход Лада Гранта по кругу получается около 7,5 л на сотню. Зимой по городу с печкой доходит до 10 л, это главный фактор, который влияет на повышение расхода топлива.
 Езжу много как в городе, так и по трассе. Расход вполне адекватный от 5,5 до 10 л.
Езжу много как в городе, так и по трассе. Расход вполне адекватный от 5,5 до 10 л.О том, как без особых усилий и затрат дать вторую жизнь автомобилю, второй ресурс, дополнительную мощность и всегда экономить около 1 литра топлива во всех режимах см по ссылке вверху любой страницы сайта.
Или в статье через строку «Поиск»: Вторая жизнь и второй ресурс для любого автомобиля, экономия топлива и дополнительная мощность. Как этого достичь?
Как этого достичь?
Всем удачи на дорогах!
Поделиться с друзьями:
Твитнуть
Поделиться
Плюсануть
Поделиться
Отправить
Класснуть
Линкануть
Запинить
Adblock
detector
Высокопроизводительное исследование FRET-детектируемого ответа ионов металлов в клетках млекопитающих
- Список журналов
- Рукописи авторов HHS
- PMC3297669
J Am Chem Soc. Авторская рукопись; доступно в PMC 2013 8 февраля.
Опубликовано в окончательной редакции как:
J Am Chem Soc. 2012 г., 8 февраля; 134(5): 2488–2491.
Published online 2012 Jan 25. doi: 10.1021/ja2101592
PMCID: PMC3297669
NIHMSID: NIHMS354211
PMID: 22260720
, † ‡ , † § , † , † ‡ * и † *
Информация об авторе Информация об авторских правах и лицензии Отказ от ответственности
- Дополнительные материалы
Генетически кодируемые датчики ионов металлов на основе FRET являются важными инструментами для изучения динамики ионов металлов в живых клетках. Мы представляем микрофлюидный проточный цитометр с временным разрешением, способный характеризовать основанный на FRET динамический отклик датчиков ионов металлов в клетках млекопитающих при пропускной способности 15 клеток/сек с временным окном, охватывающим от нескольких мс до нескольких секунд после смешивания клеток. с экзогенными лигандами. Мы использовали этот инструмент для изучения клеточной гетерогенности Zn 9. 0027 2+ и Ca 2+ сенсорных амплитуд FRET-ответа, и демонстрируют, что кластерные карты Zn 2+ сенсорных изменений FRET разрешают множественные субпопуляции. Мы также измерили кинетику реакции сенсора in vivo , вызванную изменениями концентраций Zn 2+ и Ca 2+ . Мы наблюдали ~ 30-кратную разницу между внеклеточными и внутриклеточными сенсорами.
0027 2+ и Ca 2+ сенсорных амплитуд FRET-ответа, и демонстрируют, что кластерные карты Zn 2+ сенсорных изменений FRET разрешают множественные субпопуляции. Мы также измерили кинетику реакции сенсора in vivo , вызванную изменениями концентраций Zn 2+ и Ca 2+ . Мы наблюдали ~ 30-кратную разницу между внеклеточными и внутриклеточными сенсорами.
Генетически кодируемые флуоресцентные биосенсоры произвели революцию в изучении динамических процессов в живых клетках и предоставили важную информацию о механизмах широкого спектра клеточных процессов из Ca 2+ гомеостаз активности киназы 1-3 . Теперь доступно более 120 датчиков, которые реагируют флуоресценцией на различные аналиты 3 . Эти датчики на основе FRET могут создавать зависящие от времени карты концентрации или активности ионов, малых лигандов или макромолекул в живых клетках. Для решения задачи многомерной визуализации динамический диапазон и кинетика отклика биосенсоров являются критическими атрибутами, поскольку они напрямую влияют на пространственное и временное разрешение сенсора. Кроме того, биологические измерения по своей природе неоднородны с разными уровнями межклеточных вариаций 4 . Наблюдаемая изменчивость может возникать из-за неоднородности сенсорных молекул, сообщающих об измерении, или из-за внутренней биологической изменчивости. Характеристика неоднородности населения важна: для первых она определяет чувствительность датчика и разрешающую способность; и для последнего разрешение изменчивости от клетки к клетке является важным шагом к пониманию происхождения и механизмов биологического шума.
Кроме того, биологические измерения по своей природе неоднородны с разными уровнями межклеточных вариаций 4 . Наблюдаемая изменчивость может возникать из-за неоднородности сенсорных молекул, сообщающих об измерении, или из-за внутренней биологической изменчивости. Характеристика неоднородности населения важна: для первых она определяет чувствительность датчика и разрешающую способность; и для последнего разрешение изменчивости от клетки к клетке является важным шагом к пониманию происхождения и механизмов биологического шума.
Метод, способный определять эти свойства с высокой пропускной способностью и разрешением для одной клетки, необходим для характеристики сенсоров в клеточных популяциях или клеточных библиотеках, а также для упрощения дальнейшей доработки набора инструментов для флуоресцентных сенсоров. Доступные биоаналитические методы ограничены в своих возможностях для выполнения этой задачи. Сортировка клеток с активированной флуоресценцией (FACS) 5 — это мощный высокопроизводительный метод скрининга или сортировки клеток на основе их флуоресцентных свойств. Однако измеренные сигналы отражают одиночные снимки клеточных состояний и, таким образом, дают мало информации о динамике или кинетике конкретного процесса. Автоматизированная флуоресцентная микроскопия отдельных бактериальных клеток была проведена в массивах микрожидкостных систем для изучения динамики экспрессии генов в библиотеке из 1400 штаммов 6 , однако такой подход, основанный на использовании одного члена библиотеки на микрофлюидный канал, затрудняет расширение исследования. применение к более крупным библиотекам или клеточным популяциям. Другие методы, такие как спектроскопия с остановленным потоком 7 можно использовать для измерения in vitro кинетики химической реакции с временным разрешением менее миллисекунды, но, как правило, нельзя проводить измерения на живых клетках млекопитающих (за исключением одной линии гемопоэтических клеток мыши, которая может пережить турбулентное перемешивание в смеситель с верхним потоком и временным разрешением 10 мс 8 ) и с манипуляциями с отдельными клетками (тогда как остановленный поток обеспечивает среднее значение по популяции), и, следовательно, не подходят для высокопроизводительных анализов на основе клеток.
Однако измеренные сигналы отражают одиночные снимки клеточных состояний и, таким образом, дают мало информации о динамике или кинетике конкретного процесса. Автоматизированная флуоресцентная микроскопия отдельных бактериальных клеток была проведена в массивах микрожидкостных систем для изучения динамики экспрессии генов в библиотеке из 1400 штаммов 6 , однако такой подход, основанный на использовании одного члена библиотеки на микрофлюидный канал, затрудняет расширение исследования. применение к более крупным библиотекам или клеточным популяциям. Другие методы, такие как спектроскопия с остановленным потоком 7 можно использовать для измерения in vitro кинетики химической реакции с временным разрешением менее миллисекунды, но, как правило, нельзя проводить измерения на живых клетках млекопитающих (за исключением одной линии гемопоэтических клеток мыши, которая может пережить турбулентное перемешивание в смеситель с верхним потоком и временным разрешением 10 мс 8 ) и с манипуляциями с отдельными клетками (тогда как остановленный поток обеспечивает среднее значение по популяции), и, следовательно, не подходят для высокопроизводительных анализов на основе клеток.
Здесь мы описываем микрофлюидный проточный цитометр с временным разрешением (рис. S1) для изучения изменений FRET, гетерогенности и кинетики биохимических реакций на уровне отдельных клеток. Клеточные реакции инициируются в отдельных клетках с использованием контролируемого потока жидкости в микрофлюидике 9-11 , а ход реакции отслеживается датчиками FRET. Ламинарный поток в микронном масштабе 12 и методы гидродинамической фокусировки 13 позволяют нам добиться быстрого инициирования реакции, точного управления потоком и высокопроизводительного обнаружения на уровне одной клетки. Система клапанов управления потоком () изолировала ячейки и буферные резервуары и обеспечивала непрерывное переключение буфера. Потоки клеток и буферов, управляемые давлением, втекают в микрофлюидическое устройство и встречаются на входном стыке (). На стыке клетки быстро подвергаются воздействию химических веществ, переносимых буферной линией тока, гидродинамически фокусируются в один файл и подвергаются флуоресцентному исследованию при прохождении через два лазерных луча, разделенных линией задержки различной длины. Обнаруженный сигнал представляет собой серию гауссовых импульсов, ширина импульса которых определяется временем пребывания ячейки в луче возбуждения. Чтобы сопоставить импульсы от одной и той же ячейки в этой последовательности сигналов (), мы отрегулировали скорость потока вдоль канала, используя метод двойной гидродинамической фокусировки () так, чтобы ширина импульса была разной в двух точках измерения. Используя этот метод, который «маркирует» сигналы в разных точках измерения (), мы можем извлечь время прохождения ячейки () и сопоставить пары сигналов в последовательности сигналов, когда количество ячеек (n толпа ) протекают внутри канала опроса одновременно ( и ). Эффективность сопоставления пар составляет> 85% в худшем случае (n толпа > 5) (рис. S2). Измерения проводятся в приборах с длиной линии задержки ≈ 60 мм, за исключением накладных датчиков D3pD, для которых также используются более короткие линии задержки 4 мм и 8 мм для получения временных интервалов до 1 мс.
Обнаруженный сигнал представляет собой серию гауссовых импульсов, ширина импульса которых определяется временем пребывания ячейки в луче возбуждения. Чтобы сопоставить импульсы от одной и той же ячейки в этой последовательности сигналов (), мы отрегулировали скорость потока вдоль канала, используя метод двойной гидродинамической фокусировки () так, чтобы ширина импульса была разной в двух точках измерения. Используя этот метод, который «маркирует» сигналы в разных точках измерения (), мы можем извлечь время прохождения ячейки () и сопоставить пары сигналов в последовательности сигналов, когда количество ячеек (n толпа ) протекают внутри канала опроса одновременно ( и ). Эффективность сопоставления пар составляет> 85% в худшем случае (n толпа > 5) (рис. S2). Измерения проводятся в приборах с длиной линии задержки ≈ 60 мм, за исключением накладных датчиков D3pD, для которых также используются более короткие линии задержки 4 мм и 8 мм для получения временных интервалов до 1 мс. Благодаря точному управлению потоком и гибкой конструкции устройства время прохождения ячейки можно настроить в пределах временного окна от 1 мс до 10 секунд, сохраняя при этом пропускную способность оценки FRET одной ячейки с временным разрешением. В наших типичных экспериментальных условиях, описанных во вспомогательной информации, система способна измерять одноточечный FRET при пропускной способности > 50 ячеек/с или изменение FRET отдельных ячеек при пропускной способности до 15 ячеек/с.
Благодаря точному управлению потоком и гибкой конструкции устройства время прохождения ячейки можно настроить в пределах временного окна от 1 мс до 10 секунд, сохраняя при этом пропускную способность оценки FRET одной ячейки с временным разрешением. В наших типичных экспериментальных условиях, описанных во вспомогательной информации, система способна измерять одноточечный FRET при пропускной способности > 50 ячеек/с или изменение FRET отдельных ячеек при пропускной способности до 15 ячеек/с.
Открыть в отдельном окне
Экспериментальная установка высокопроизводительной микрофлюидной платформы для динамического измерения FRET в живых клетках. (a) Схема возбуждения/обнаружения для измерения FRET в микрофлюидике. (b) Репрезентативная конструкция канала микрофлюидного устройства (длина линии задержки ~ 60 мм). Несколько ячеек (обозначенных красными точками, размер не в масштабе) могут одновременно проходить внутри канала опроса. (c) Система клапана регулирования расхода. (d) Изображение микрофлюидного потока на устройстве. Стрелки показывают положение входа в клеточный буфер и точки измерения флуоресценции.
(d) Изображение микрофлюидного потока на устройстве. Стрелки показывают положение входа в клеточный буфер и точки измерения флуоресценции.
Открыть в отдельном окне
Анализ данных и алгоритм сопоставления пар. (a) Последовательность сигналов, генерируемых флуоресцентными клетками, пересекающими возбуждающие лазерные лучи. (b) Распределение ширины импульса (стандартное отклонение гауссовой аппроксимации) в двух точках измерения; разная ширина обусловлена разной скоростью потока, создаваемой методом двойной гидродинамической фокусировки. (c, d) Гистограмма межимпульсного интервала извлекается из последовательности импульсов путем предположения пробного n скопления и подсчета временных интервалов, попадающих в разные интервалы времени. Время прохождения ячейки получается из гистограммы. (c) Короткое время в пути при высоком давлении 90–105 . (d) Большое время в пути при низком давлении.
Мы продемонстрировали полезность этого метода для двух приложений: количественный скрининг межклеточной неоднородности и измерение мс с временным разрешением in vivo кинетика. Для первого применения этот метод позволяет инициировать клеточные реакции в течение нескольких мс, измерять изменения отношения FRET с разрешением одной клетки и анализировать изменчивость FRET на уровне популяции. Для второго приложения наш прибор позволяет точно контролировать микрофлюидный поток для измерения кинетики отклика датчиков в их естественной клеточной среде во временных окнах от 1 мс до 10 секунд. Этот промежуток времени является важной физиологически значимой областью, в которой протекают многочисленные биологические реакции, т.е. фолдинг белка и эндоцитоз синаптических пузырьков 14,15 , бывает. Однако это особенно трудное временное окно для измерения кинетики клеточных процессов с помощью флуоресцентной микроскопии, потому что относительно медленная макроскопическая диффузия и перемешивание часто происходят в сравнимых временных масштабах и, следовательно, могут значительно влиять на измеряемую динамику. Здесь мы используем методы микрофлюидики, чтобы сократить время перемешивания, контролируемого диффузией, до нескольких мс.
Здесь мы используем методы микрофлюидики, чтобы сократить время перемешивания, контролируемого диффузией, до нескольких мс.
Время диффузии находится в квадратичной зависимости от расстояния. Следовательно, с помощью методов гидродинамической фокусировки 13 , в котором поток, содержащий клетки, сжимается до «струи» 10 мкм двумя буферными растворами, содержащими аналит, время достижения молекулами клеток может быть значительно сокращено. На рис. S3 показано смоделированное поле концентрации Ca 2+ в виде двумерного изображения поверхности и одномерного графика поперечного сечения. Профиль концентрации по сечению показывает, что Ca 2+ быстро проникает в центр гидродинамического очага, достигая ~3 мМ через ~2,5 мс после входа. Эта концентрация примерно в 1000 раз превышает минимальную точку насыщения металлических сенсоров, которые имеют константы диссоциации в диапазоне от пМ до мкМ.
Мы измерили кинетику ответа FRET и гетерогенность ответа клеток HeLaS, экспрессирующих внутриклеточные сенсоры Ca 2+ и Zn 2+ . В частности, мы использовали генетически кодируемые сенсоры Ca 2+ (D3cpV 16 , экспрессированные в цитозоле и D3pd, экспонированные на внеклеточной поверхности) и один сенсор Zn 2+ (ZapCY1 17 , экспрессированные в цитозоле). . Эти датчики сообщают об изменении концентрации внутри- или внеклеточных ионов металлов путем рекрутирования одного (или нескольких) ионов металла в свои металлосвязывающие домены, вызывая конформационные изменения, ведущие к более высокой эффективности FRET между донором (CFP) и акцептор (YFP) (рис. S4).
В частности, мы использовали генетически кодируемые сенсоры Ca 2+ (D3cpV 16 , экспрессированные в цитозоле и D3pd, экспонированные на внеклеточной поверхности) и один сенсор Zn 2+ (ZapCY1 17 , экспрессированные в цитозоле). . Эти датчики сообщают об изменении концентрации внутри- или внеклеточных ионов металлов путем рекрутирования одного (или нескольких) ионов металла в свои металлосвязывающие домены, вызывая конформационные изменения, ведущие к более высокой эффективности FRET между донором (CFP) и акцептор (YFP) (рис. S4).
Сначала мы подтвердили, что реакции связывания металлов могут индуцироваться микрофлюидным потоком и наблюдаться как изменения FRET. Перед микрофлюидным экспериментом клетки предварительно обрабатывали хелаторами металлов, чтобы истощить сенсоры целевых ионов металлов (, например, EGTA для Ca 2+ или TPEN для Zn 2+ ), таким образом поддерживая сенсоры в безметалловое состояние с минимальным коэффициентом FRET. В контрольных экспериментах клетки не показали изменений в своих логометрических значениях FRET при смешивании с не содержащими металлов буферными потоками, содержащими хелаторы (рис. S5). Кроме того, в популяцию клеток D3cpV вводят флуоресцентные гранулы (не реагирующие на возмущение ионами металлов), которые служат внутренней калибровкой реакции. При измерении отклика буферные потоки содержат ионы металлов, что приводит к воздействию на клетки в течение мс от входного соединения. Как и ожидалось от конструкции датчика, клетки демонстрируют значительное увеличение FRET после временной задержки > 1 с (1).
В контрольных экспериментах клетки не показали изменений в своих логометрических значениях FRET при смешивании с не содержащими металлов буферными потоками, содержащими хелаторы (рис. S5). Кроме того, в популяцию клеток D3cpV вводят флуоресцентные гранулы (не реагирующие на возмущение ионами металлов), которые служат внутренней калибровкой реакции. При измерении отклика буферные потоки содержат ионы металлов, что приводит к воздействию на клетки в течение мс от входного соединения. Как и ожидалось от конструкции датчика, клетки демонстрируют значительное увеличение FRET после временной задержки > 1 с (1).
Открыть в отдельном окне
Ca 2+ и Zn 2+ — сенсорные ответы в микрофлюидике. (a) Ca 2+ ответ клеток HeLa, экспрессирующих сенсор D3cpV Ca 2+ при T = 1,29 ± 0,06 с, запускаемый буфером, нагруженным Ca 2+ , содержащим 30 мМ Ca 2+ и 49 мкМ иономицин. (●) пятно 1, () пятно 2. (b) Карта кластера FRET (FRET в пятне 2 по сравнению с FRET в пятне 1) клеток и гранул D3cpV-HeLa. (в) Zn 2+ ответ клеток HeLa, экспрессирующих датчик ZapCY1 Zn 2+ при T = 3,43 ± 0,14 с, запускаемый буфером, нагруженным Zn 2+ , содержащим 4 мМ Zn 2+ и 70 мкМ пиритиона. (●) точка 1, () точка 2. (d) Карта кластера FRET разрешает четыре субпопуляции (p1-p4).
(в) Zn 2+ ответ клеток HeLa, экспрессирующих датчик ZapCY1 Zn 2+ при T = 3,43 ± 0,14 с, запускаемый буфером, нагруженным Zn 2+ , содержащим 4 мМ Zn 2+ и 70 мкМ пиритиона. (●) точка 1, () точка 2. (d) Карта кластера FRET разрешает четыре субпопуляции (p1-p4).
Пропускная способность измерения изменения FRET на основе одной ячейки определяется эффективностью метода сопоставления пар и n толпы и варьируется от 15 ячеек в секунду для коротких окон времени (< 0,5 секунды) до ~1 ячейка/секунда для временных окон продолжительностью более 5 секунд. Статистическая информация, такая как среднее значение, стандартное отклонение или асимметрия (асимметрия распределения вероятностей) ответа FRET для тысяч клеток, может быть получена в течение нескольких минут. По сравнению с коэффициентом вариации (CV шариков = 0,07 ± 0,02 ) шариков с примесями, CV клеток больше (CV клеток > 0,1), что указывает на присущее разнообразие сенсорной экспрессии и отношения FRET в живых клетках. Ранее было замечено, что генетически закодированные сенсоры, встроенные в живые клетки, демонстрируют межклеточную изменчивость 18 . Неоднородность также зависит от состояния металлического связывания датчиков. Как показано и наблюдается как общая тенденция, FRET в точке 2 (Ca 2+ или Zn 2+ -связанный и высокий коэффициент FRET) демонстрирует больший CV по сравнению с пятном 1 (Ca 2+ или Zn 2+ -свободный и низкий коэффициент FRET). Это наблюдение предполагает, что эффективность FRET в связанном с металлом «активном» состоянии, вероятно, более чувствительна к небольшим межклеточным вариациям сенсорных молекул (, например, конформационная гетерогенность, посттрансляционные модификации, термодинамический шум, и т. д. ). , или клеточные свойства ( например размер ячейки, стадия, металлическая буферная емкость, и т. д. ).
Ранее было замечено, что генетически закодированные сенсоры, встроенные в живые клетки, демонстрируют межклеточную изменчивость 18 . Неоднородность также зависит от состояния металлического связывания датчиков. Как показано и наблюдается как общая тенденция, FRET в точке 2 (Ca 2+ или Zn 2+ -связанный и высокий коэффициент FRET) демонстрирует больший CV по сравнению с пятном 1 (Ca 2+ или Zn 2+ -свободный и низкий коэффициент FRET). Это наблюдение предполагает, что эффективность FRET в связанном с металлом «активном» состоянии, вероятно, более чувствительна к небольшим межклеточным вариациям сенсорных молекул (, например, конформационная гетерогенность, посттрансляционные модификации, термодинамический шум, и т. д. ). , или клеточные свойства ( например размер ячейки, стадия, металлическая буферная емкость, и т. д. ).
Два генетически закодированных сенсора, исследованных здесь, демонстрируют совершенно разные профили гетерогенности. В отличие от примерно унимодального распределения FRET D3cpV, измерение FRET ZapCY1 разрешает четыре субпопуляции с различиями либо в статическом значении FRET, либо в динамическом ответе FRET (4). Статистический анализ ответа Zn 2+ 3358 клеток из различных измерений неизменно показывает, что субпопуляция P 2 (35,2% ± 3,1 %) демонстрирует ожидаемый Zn 2+ -индуцированный ответ, в то время как остальные три субпопуляции (P 1 (4,8 % ± 1,5 %), P 3 (20,2 % ± 2,6 %). ) и P 4 (39,8% ± 2,2%)) демонстрируют незначительный или слабый отклик Zn 2+ (). Внутри отдельных субпопуляций также существует изменчивость в распределении. Три «невосприимчивые» субпопуляции демонстрируют низкие, высокие и средние исходные значения FRET, что позволяет предположить, что часть этой гетерогенности может возникать из-за протеолиза или сенсорной нестабильности в клеточной среде. Например, Р 4 демонстрирует низкую флуоресценцию CFP, что приводит к высокому коэффициенту FRET, а P 1 имеет низкую флуоресценцию YFP, что приводит к низкому коэффициенту FRET.
В отличие от примерно унимодального распределения FRET D3cpV, измерение FRET ZapCY1 разрешает четыре субпопуляции с различиями либо в статическом значении FRET, либо в динамическом ответе FRET (4). Статистический анализ ответа Zn 2+ 3358 клеток из различных измерений неизменно показывает, что субпопуляция P 2 (35,2% ± 3,1 %) демонстрирует ожидаемый Zn 2+ -индуцированный ответ, в то время как остальные три субпопуляции (P 1 (4,8 % ± 1,5 %), P 3 (20,2 % ± 2,6 %). ) и P 4 (39,8% ± 2,2%)) демонстрируют незначительный или слабый отклик Zn 2+ (). Внутри отдельных субпопуляций также существует изменчивость в распределении. Три «невосприимчивые» субпопуляции демонстрируют низкие, высокие и средние исходные значения FRET, что позволяет предположить, что часть этой гетерогенности может возникать из-за протеолиза или сенсорной нестабильности в клеточной среде. Например, Р 4 демонстрирует низкую флуоресценцию CFP, что приводит к высокому коэффициенту FRET, а P 1 имеет низкую флуоресценцию YFP, что приводит к низкому коэффициенту FRET. Важно отметить, что эта гетерогенность не была выявлена в экспериментах по микроскопии отдельных клеток, вероятно, из-за сравнительно низкой производительности микроскопии и субъективного характера выбора «отзывчивых клеток»; для получения набора данных, эквивалентного представленному в, потребуется не менее 35 часов на стандартном флуоресцентном микроскопе, предполагая щедрую оценку 10 трансфицированных клеток в поле зрения и ~ 1 час на калибровочный эксперимент.
Важно отметить, что эта гетерогенность не была выявлена в экспериментах по микроскопии отдельных клеток, вероятно, из-за сравнительно низкой производительности микроскопии и субъективного характера выбора «отзывчивых клеток»; для получения набора данных, эквивалентного представленному в, потребуется не менее 35 часов на стандартном флуоресцентном микроскопе, предполагая щедрую оценку 10 трансфицированных клеток в поле зрения и ~ 1 час на калибровочный эксперимент.
Измерения на основе популяции ясно показывают, что функции распределения, описывающие реакцию FRET, могут принимать различные формы, либо в виде одномодального распределения FRET, либо в виде мультимодального распределения с отдельными субпопуляциями. Кроме того, разная вариабельность FRET связана с каждым состоянием сенсора (исходным и активным) и с каждой субпопуляцией (p1-p4). Наши данные показывают, что два датчика FRET со схожими молекулярными конструкциями приводят к существенно разным профилям неоднородности. Способность идентифицировать и определять эту неоднородность имеет важные последствия для конструкции и использования датчика.
Способность идентифицировать и определять эту неоднородность имеет важные последствия для конструкции и использования датчика.
Недавние разработки в области дизайна биосенсоров привели к значительному повышению чувствительности сенсоров на основе FRET 17,19,20 , однако характеристика и оптимизация их in vivo кинетических свойств отстает. Хорошо охарактеризованная, быстрая кинетика отклика сенсора имеет решающее значение для исследований клеточной динамики, таких как измерения нервной активности 21,22 . Оптимальный датчик должен достоверно сообщать о кинетике биологического сигнала; медленно реагирующий датчик может исказить или даже пропустить переходный сигнал. Пользуясь преимуществом управления микрофлюидным потоком и измерениями FRET с временным разрешением, мы расширяем этот метод микрофлюидики для исследования кинетики датчиков в клеточной среде. Кинетика измеряется как среднее значение FRET нормальной популяции, реагирующей на ионы металлов, и наносится на график как функция времени прохождения клеток. Реакция с временным разрешением генетически кодируемого Ca 9Датчики 0027 2+ и Zn 2+ в клетках HeLa (и рис. S6) варьируются от ~ 20 мс (внеклеточно отображаемый сенсор) до нескольких секунд (внутриклеточно экспрессируемый сенсор).
Реакция с временным разрешением генетически кодируемого Ca 9Датчики 0027 2+ и Zn 2+ в клетках HeLa (и рис. S6) варьируются от ~ 20 мс (внеклеточно отображаемый сенсор) до нескольких секунд (внутриклеточно экспрессируемый сенсор).
Открыть в отдельном окне
Кинетика датчиков в ячейках Hela с временным разрешением. (a) Активация внеклеточного сенсора (D3pd). (b) D3pd показывает наблюдаемую скорость k obs =43 ± 6 с -1 при воздействии 30 мМ Ca 2+ . (c) Активация внутриклеточного сенсора (D3cpv). (d) D3cpV показывает наблюдаемую скорость ответа k obs =1,4 ± 0,2 с -1 при воздействии 30 мМ Ca 2+ и 49 мкМ иономицина. ( — экспериментальные данные, — — экспоненциальная аппроксимация кривой.
Сенсоры, экспрессируемые в разных клеточных локациях, вероятно, демонстрируют различную кинетику реакции. Для сенсоров, отображаемых на внеклеточной мембране (), измеренная константа скорости в значительной степени определяется собственной скоростью включения/выключения сенсоров и применяемой концентрацией ионов металлов. Для внутриклеточных экспрессируемых сенсоров (1) наблюдаемая кинетика дополнительно ограничивается переносом ионов металлов через плазматическую мембрану. Чтобы изучить этот эффект, мы экспрессировали сенсоры в разных местах клеток и сравнивали скорость их отклика в идентичных экспериментальных условиях. Наблюдаемая константа скорости D3pd (поверхность, отображающая Ca 2+ датчик) is k наб ≈ 43 ± 6 с −1 (). Напротив, внутриклеточный сенсор D3cpV Ca 2+ аналогичной конструкции демонстрирует кинетику ответа примерно в 30 раз медленнее (k ≈ 1,4 ± 0,2 с -1 ) (). Различные временные масштабы указывают на то, что транспорт ионов металлов через клеточную мембрану, вероятно, является стадией, ограничивающей скорость активации внутриклеточного сенсора.
Для сенсоров, отображаемых на внеклеточной мембране (), измеренная константа скорости в значительной степени определяется собственной скоростью включения/выключения сенсоров и применяемой концентрацией ионов металлов. Для внутриклеточных экспрессируемых сенсоров (1) наблюдаемая кинетика дополнительно ограничивается переносом ионов металлов через плазматическую мембрану. Чтобы изучить этот эффект, мы экспрессировали сенсоры в разных местах клеток и сравнивали скорость их отклика в идентичных экспериментальных условиях. Наблюдаемая константа скорости D3pd (поверхность, отображающая Ca 2+ датчик) is k наб ≈ 43 ± 6 с −1 (). Напротив, внутриклеточный сенсор D3cpV Ca 2+ аналогичной конструкции демонстрирует кинетику ответа примерно в 30 раз медленнее (k ≈ 1,4 ± 0,2 с -1 ) (). Различные временные масштабы указывают на то, что транспорт ионов металлов через клеточную мембрану, вероятно, является стадией, ограничивающей скорость активации внутриклеточного сенсора.
В целом мы представляем новый подход к измерению FRET одной ячейки с временным разрешением, который является гибким и высокопроизводительным. Этот метод представляет собой обобщение проточной цитометрии с временным разрешением путем добавления измерения времени к измерениям FRET с одной клеткой. Эта система способна контролировать и возмущать внеклеточную среду, а также оптически исследовать кинетику клеточных реакций. Мы разрешили значительно отличающиеся профили гетерогенности, связанные с различными генетически кодируемыми сенсорами, и сообщили о кинетике сенсоров в разных клеточных местах, раскрывая как присущую сенсорам кинетику, так и кинетику переноса ионов через мембрану. Эти измерения были бы невозможны при использовании традиционной флуоресцентной микроскопии, проточной цитометрии или спектроскопии с остановленным потоком. Наш метод может быть легко расширен для крупномасштабного скрининга на основе кинетики или измерения других оптически меченых молекулярных взаимодействий или реакций в живых клетках.
1_si_001
Нажмите здесь для просмотра. (419K, pdf)
Мы хотели бы отметить финансовую поддержку начального гранта CU-NIST и гранта GM083849 NIH для A.E.P. и RJ, и GM084027 в AEP, а также из Физического пограничного центра JILA (в RJ). Р.Дж. является сотрудником отдела квантовой физики NIST. П.Дж.Д. был поддержан грантом на обучение биофизике (T32 GM-065103).
ДОПОЛНИТЕЛЬНОЕ СОДЕРЖИМОЕ
Дополнительная информация . Опорные рисунки, материалы и методы. Этот материал доступен бесплатно в Интернете по адресу http://pubs.acs.org».
Вклад авторов
Эта рукопись была написана благодаря вкладу всех авторов. Все авторы одобрили окончательный вариант рукописи.
(1) Giepmans BNG, Adams SR, Ellisman MH, Tsien RY. Наука. 2006; 312: 217–224. [PubMed] [Google Scholar]
(2) Палмер А.Е., Цинь И., Парк Дж.Г., МакКомбс Дж.Е. Тенденции биотехнологии. 2011;29: 144–152. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
(3) Newman RH, Fosbrink MD. Чжан, J. Chem. Rev. (Вашингтон, округ Колумбия, США) 2011; 111:3614–3666. [Бесплатная статья PMC] [PubMed] [Google Scholar]
(4) Lidstrom ME, Konopka MC. Нац. хим. биол. 2010;6:705–712. [PubMed] [Google Scholar]
(5) Bonner WA, Hulett HR, Sweet RG, Herzenberg LA. преподобный наук. Инструм. 1972; 43: 404–409. [PubMed] [Google Scholar]
(6) Танигути Ю., Чой П.Дж., Ли Г.В., Чен Х.И., Бабу М., Хирн Дж., Эмили А., Се Х.С. Наука. 2010;329: 533–538. [Бесплатная статья PMC] [PubMed] [Google Scholar]
(7) Chance B. Rev. Sci. Инструм. 1951; 22: 619–627. [Google Scholar]
(8) Wilkinson JC, Beechem JM, Staros JVJ. Прием Сигнал Преобразования Res. 2002; 22: 357–371. [PubMed] [Google Scholar]
(9) Whitesides GM. Природа. 2006; 442: 368–373. [PubMed] [Google Scholar]
(10) Squires TM, Quake SR. Преподобный Мод. физ. 2005; 77: 977–1026. [Google Scholar]
(11) Beebe DJ, Mensing GA, Walker GM. Анну. Преподобный Биомед. англ. 2002; 4: 261–286. [PubMed] [Академия Google]
Анну. Преподобный Биомед. англ. 2002; 4: 261–286. [PubMed] [Академия Google]
(12) Константинеску В.Н. Ламинарное вязкое течение. Спрингер-Верлаг; Нью-Йорк: 1995. [Google Scholar]
(13) Найт Дж. Б., Вишванат А., Броди Дж. П., Остин Р. Х. физ. Преподобный Летт. 1998;80 [Google Scholar]
(14) McCammon JA, Harvey SC. Динамика белков и нуклеиновых кислот Кембриджского унив. Нажимать. 1987 [Google Scholar]
(15) Ryan TA, Smith SJ, Reuter H. Proc. Натл. акад. науч. США, 1996; 93:5567–5571. [Бесплатная статья PMC] [PubMed] [Google Scholar]
(16) Палмер А.Е., Джакомелло М., Кортемме Т., Хайрес С.А., Лев-Рам В., Бейкер Д., Циен Р.Ю. хим. биол. 2006; 13: 521–530. [PubMed] [Академия Google]
(17) Цинь Ю., Диттмер П.Дж., Парк Дж.Г., Янсен К.Б., Палмер А.Е. проц. Натл. акад. науч. США 2011;108:7351–7356. [Бесплатная статья PMC] [PubMed] [Google Scholar]
(18) Dittmer PJ, Miranda JG, Gorski JA, Palmer AE. Дж. Биол. хим. 2009; 284:16289–16297. [Бесплатная статья PMC] [PubMed] [Google Scholar]
(19) Horikawa K, Yamada Y, Matsuda T, Kobayashi K, Hashimoto M, Matsu-ura T, Miyawaki A, Michikawa T, Mikoshiba K, Nagai T. Nat . Методы. 2010;7:729–U88. [PubMed] [Академия Google]
Nat . Методы. 2010;7:729–U88. [PubMed] [Академия Google]
(20) Манк М., Райфф д. F., Heim N, Friedrich MW, Borst A, Gries-beck O. Biophys. Дж. 2006; 90:1790–1796. [Бесплатная статья PMC] [PubMed] [Google Scholar]
(21) Wallace DJ, Borgloh S, Astori S, Yang Y, Bausen M, Kugler S, Palmer AE, Tsien RY, Sprengel R, Kerr JND, Denk W, Хасан МТ. Нац. Методы. 2008; 5: 797–804. [PubMed] [Google Scholar]
(22) Tian L, Hires SA, Mao T, Huber D, Chiappe ME, Chalasani SH, Petreanu L, Akerboom J, McKinney SA, Schreiter ER, Bargmann CI, Jayaraman V, Svoboda K , Лугер ЛЛ. Нац. Методы. 2009 г.;6:875–U113. [Бесплатная статья PMC] [PubMed] [Google Scholar]
KBLEE Group – Институт нанобиомедицинских исследований
Добро пожаловать на сайт KBLee Group!
Последние новости
- Сара получает стипендию программы обучения биотехнологии, поздравляем!
- Этой осенью д-р Джинхо Юн приступит к работе в Католическом университете Кореи (Корея) в качестве доцента. Поздравляю!
- Профессор Ки-Бум Ли получил звание заслуженного профессора. Поздравляю!
Профессор Ли хотел бы поблагодарить группу KBLEE и сотрудников за их поддержку и вклад. - Брайан успешно защитил кандидатскую диссертацию. диссертация. Поздравляем, доктор Конли!
- Брэндон получил стипендию по обучению переводу нейробиологии в терапию, поздравляем!
- Мероприятие T32 Immersion 2022 года пройдет 14–15 января с 10:00 до 18:00. Для получения дополнительной информации, включая ссылку для регистрации, см. вкладку «События» .
- Д-р Джин-Ха Чой поступил на работу в Чонбукский национальный университет (Корея) в качестве доцента.
Поздравляю! - Профессор Ки-Бум Ли стал лауреатом премии Grossman Innovation Prize 2021, поздравляем!
- Брайан получил премию Рейда 2020 года, поздравляем!
- Джефф получил премию Ван Дайка 2020 года, поздравляем!
- Худифа выиграла премию Рейда 2019 года, поздравляем!
- Крис выиграл премию Рейда 2018 года, поздравляем!
- Худифа выиграла премию Ван Дайка 2018 года, поздравляем!
- Top получил премию Ван Дайка 2018 года, поздравляем!
- Амин выиграл первую ежегодную премию BME за лидерство, поздравляем!
- Наша работа над нанокаркасом MnO 2 теперь во многих средствах массовой информации!
Eurekalert, R&D Magazine, Innovators Magazine, Drug Target Review, The Politico Post, Science Daily, Phys. Org, Innovations Report, Jersey Tribune, News-Medical, Bioengineer, лента медицинских новостей, (e) Science News, Front Line Genomics , Science Bulletin, BioSpectrum Asia Edition, W3Live News, BioNews Central, Dove Med, Career Biotech, ChemEurope - Крис выиграл стипендию Центра исследований спинного мозга в Нью-Джерси, поздравляем!
- Skylar получает стипендию программы обучения биотехнологии!
- Худифа, Летао и Исяо выиграли Фонд профессионального развития 2018 года для преподавателей и ассистентов выпускников!
 Поздравляю!
Поздравляю!  Org, Innovations Report, Jersey Tribune, News-Medical, Bioengineer, лента медицинских новостей, (e) Science News, Front Line Genomics , Science Bulletin, BioSpectrum Asia Edition, W3Live News, BioNews Central, Dove Med, Career Biotech, ChemEurope
Org, Innovations Report, Jersey Tribune, News-Medical, Bioengineer, лента медицинских новостей, (e) Science News, Front Line Genomics , Science Bulletin, BioSpectrum Asia Edition, W3Live News, BioNews Central, Dove Med, Career Biotech, ChemEurope- Рукопись Джеффа, Топа, Брайана, Джинхо и Яннана, опубликованная в Bioactive Materials: «Инъекционный биоортогональный гидрогель (BIOGEL) ускоряет регенерацию тканей в дегенерированных межпозвонковых дисках», Bioactive Materials, 2022.
Это совместная работа с профессором Инбо Ханом из Медицинской школы Университета Ча. - Рукопись Брэндона, Брайана, Яннана и Мейзи, опубликованная в Advanced Drug Delivery Reviews: «Advanced Theragnostic for the Central Nervous System (CNS) and Neurological Disorders Using Functional Inorganic Nanomaterials», Advanced Drug Delivery Reviews, 2022.
- Рукопись Летао, Брайана, Джинхо, Криса, Топа, Брэндона и Яннана, опубликованная в журнале Research: «Скрининг высокого содержания и анализ нейронных интерфейсов, полученных из стволовых клеток, с использованием комбинаторного нанотехнологического подхода и интегрированного машинного обучения», Research, 2022
- Рукопись Джеффа, опубликованная в ACS Applied Materials & Interfaces: «Усовершенствованная доставка лекарств с помощью гибридных нановолокон для улучшения дифференцировки стволовых клеток», ACS Applied Materials & Interfaces, 2022.
- Рукопись Мердада и Мейзи, опубликованная в Journal of Molecular Liquids: «Оценка антибактериальных и механических свойств стоматологических адгезивов, содержащих коллоидные наночастицы золота», Journal of Molecular Liquids, 2022.
- Рукопись Скайлара, Брэндона, Джоша и Джорджа, опубликованная в Nano Convergence: «Иммуноинженерные подходы с использованием нанотехнологий для продвижения терапевтических приложений», Nano Convergence, 2022.
- Рукопись Джинхо и Брайана, опубликованная в ACS Nano: «Сверхчувствительное электрохимическое обнаружение мутировавших вирусных РНК с разрешением по одному нуклеотиду с использованием массива нанопористых электродов (NPEA)», ACS Nano, 2022.
Это совместная работа с профессором Чон- Ву Чой из Университета Соган. - Рукопись Летао, Брайана, Топа и Брэндона, опубликованная в ACS Nano: «Прогностическое картирование биофизических сигналов для клеточной дифференцировки и перепрограммирования с использованием комбинаторных наночипов», ACS Nano, 2022.
- 4
3 90, Chris Letao
3
3 и рукопись Яннана, опубликованная в журнале Small: «Использование терапевтического потенциала внеклеточных везикул для биомедицинских применений с использованием многофункциональных магнитных наноматериалов», Small, 2022.
Это совместная работа с профессором Химин Кан из Корейского университета. - Рукопись Джейсона и Летао опубликована в National Science Review: «3D Nano-Hybrid Stem Cell Gel for Rapid Regeneration Cartilage Regeneration», National Science Review, 2022. Университет.
- Рукопись Джинхо Юна, опубликованная в журнале Small Methods: «Биомолекулярный электронный регулятор, состоящий из нанобиогибрида с электрически высвобождаемым комплексом для пространственно-временного контроля дифференцировки нейронов», Small Methods, 2021.
Это совместная работа с профессором Чон-Ву Чой из Университета Соган. - Рукопись Криса и Летао принята в журнал Science Advances: «Hybrid SMART Spheroids для усовершенствованной заместительной терапии на основе стволовых клеток», Science Advances, 2021 .
- Рукопись СангА, Исяо, Криса и Топа опубликована в Advanced Materials: «Bioengineering Approaches for the Advanced Organoid Research», Advanced Materials, 2021 .
- Рукопись Джин-Ха Чой, Летао, Брайана и Джинхо Юна, опубликованная в ACS Nano: «CRISPR-Cas12a-опосредованное обнаружение без амплификации мультивирусных ДНК с использованием SERS-активного массива наноцветков оксида графена/треугольника Au» ”, ACS Nano, 2021.
Это совместная работа с профессором Чон-Ву Чой из Университета Соган. - Наша рукопись опубликована в журнале Small: «Магнитный контроль вертикального положения иерархической наносборки лигандов регулирует дифференцировку стволовых клеток, опосредованную механосенсорным восприятием»», Small, 2021.
Это совместная работа с профессором Химин Кан из Корейского университета. - Collaboration Manuscript with Prof. Heemin Kang published in Advanced Materials : “Remote Control of Time Regulated Stretching of Ligand-Presenting Nanocoils In Situ Regulates Cyclic Адгезия и дифференцировка стволовых клеток», Advanced Materials , 2021 .
- Collaboration Manuscript with Prof. Heemin Kang accepted in Advanced Functional Materials : “Remote Switching of Elastic Movement of Decorated Ligand Nanostructures Controls the Adhesion-Regulated Polarization макрофагов-хозяев», Продвинутый Функциональный Материалы , 2021 .
- Collaboration Manuscript with Prof. Li Cai published in Experimental Neurology : “Nkx6 Promotes Neurogenesis and Attenuates Glial Scar Formation in the Injured Spinal Cord”, Экспериментальная неврология , 2021 .
- Collaboration Manuscript with Prof. Li Cai published in Molecular Therapy : “Gsx1 Promotes Locomotor Functional Recovery After Spinal Cord Injury”, Molecular Therapy , 2021 .
- Совместная рукопись с профессором Мортезой Махмуди опубликована в Molecular Pharmaceutics : “Restoring endogenous repair mechanisms to heal chronic wounds with a multi-fuctional wound dressing”, Molecular Pharmaceutics , 2021 .
- Рукопись Летао, Брайана и Топа, опубликованная в Advanced Materials: «Efffective Modulation of CNS Inhibitor Microvironment using Bio-inspired Hybrid Nanoscaffold-based Therapeutic Interventions», Advanced Materials, 2020 .
- Летао, Тэ-Хён, Хён-Йол, Джефф, Чон-Мин, Дин, Яннан и Перри. Конъюгаты для специфической для рака мультимодальная визуализация и комбинированная Therapeutics «, Усовершенствованные функциональные материалы , 2020.
- Jeff’s Collaborport Manuscript Offindable Sporrancemance Sprevancemance Pransultable Sprevancemplable. 3D-культура клеток и гистология», Advanced Materials, 2020 .
This is a collaboration work with Prof. Howon Lee at Rutgers University - Jin-Ha, Letao, and Brian’s Manuscript published in Nano Letters : “In Situ Detection of Neurotransmitters из нейронного интерфейса, полученного из стволовых клеток, на уровне отдельных клеток с помощью графен-гибридного SERS Nanobiosensing», Nano Letters, 2020
- Yixiao и рукопись Hudifah, опубликованная в ACS Applied Materials & Interfaces : «NIR-опосредованные пространственно-времен
- Jin-Ho, Jeff, and Dean’s Review Статья опубликована в Nanoscale: «Функциональные наночипы для исследования судьбы и функции стволовых клеток», Nanoscale, 2020
- Рукопись Летао, Джин-Хо, Криса и Яннана, опубликованная в Nano Letters: «Комбинаторный графен-золотогибридный SERS-наночип для двойного усиления обнаружения нейронных генов», Nano Letters, 2019
- 4 3 Chris’s 9s Обзорная статья принята в Frontiers in Cellular Neuroscience: «Bio-Inspired Nanomaterials for Delivery of Therapeutics to Лечение потери слуха», Frontiers in Cellular Neuroscience, 2019
- Рукопись Джин-Хо и Джин-Ха опубликована в ACS Nano: « Неразрушающая характеристика нейрогенеза стволовых клеток с помощью обнаружения экзосомальных миРНК на основе магнитоплазмонных наноматериалов», ACS Nano, 2019
- Jason and Letao’s Manuscript published in Biomaterials: “Programmed Degradation of a Hierarchical Nanoparticle with Redox and Light Responsivity for Self-activated Photo-chemical Enhanced Chemodynamic Therapy”, Biomaterials, 2019
- Hyeon-Yeol’s Совместная рукопись опубликована в журнале Bioconjugate Chemistry: «Включение дитиолана, содержащего аминокислоту, в белки для конкретного сайта», Bioconjugate Chemistry, 2019 г.





 3D-культура клеток и гистология», Advanced Materials, 2020 .
3D-культура клеток и гистология», Advanced Materials, 2020 . 